The Function Of Cytidine Coenzymes. Part 2
Description
This section is from the book "The Scientific Contributions Of The Ben May Laboratory For Cancer Research", by The University of Chicago. Also available from Amazon: The Scientific Contributions Of The Ben May Laboratory For Cancer Research.
The Function Of Cytidine Coenzymes. Part 2
Experimental
Enzymatic Incorporation of P-choline into Lecithin-When homogenates of rat liver were incubated with P-choline-P82 in the presence of MgClj, phosphate buffer, and a preparation of amorphous ATP (Pabst lot No. 116), a considerable incorporation of radioactivity into phospholipide took place. In confirmation of the work of Kornberg and Pricer (1) and of Rodbell and Hanahan (4), it was found that P-choline-1,2-CM was incorporated at a rate identical with that of P-choline-P32, suggesting that P-choline is incorporated as a unit into the phospholipide structure.
Table I. Incorporation Of P-Choline-P" Into Lecithin In Subcellular Fractions Of Rat Liver
Tube No. | P-choline incorporated into lipide | |
mµmoles | ||
1 | Whole homogenate, phosphatidic acid added | 5.8 |
2 | " " " " omitted | 6.3 |
3 | Particles, phosphatidic acid added | 20.0 |
4 | " " " omitted | 18.5 |
5 | Supernatant fraction, phosphatidic acid added | 2.7 |
6 | " " " " omitted | 2.6 |
Each tube contained 20 µmoles of MgCl2, 100 µmoles of phosphate buffer of pH 7.4, 10 µmoles of P-choline-P'1 (34,500 c.p.m. per micromole), and enzyme derived from approximately 0.2 gm. wet weight of rat liver. 10 pinoles of amorphous ATP (lot No. 116) were added at the beginning of the experiment, and 1.0 pinole was added every 10 minutes during the course of the reaction. 0.20 µmoles of synthetic dipalmitoyl-L-a-glycerophosphate was added as indicated. The final volume of the system was 2.0 ml. The tubes were incubated at 37° for 1 hour.
Further experiments revealed that the particulate fraction of rat fiver was considerably more active in the incorporation of P-choline into lipide than the equivalent amount of whole homogenate from which the particles were derived (Table I). For this reason, isolated particles or enzymes derived from particles were used in most subsequent experiments.
The addition of synthetic phosphatidic acid was without effect on the enzyme system.
Identification of Radioactive Product As Lecithin-In the experiments of Kornberg and Pricer (1), the product derived from P-choline was shown to be a lipide and to contain P32 and C14 in a ratio closely similar to that of the original labeled P-choline, but was not further identified. Rodbell and Hanahan (4) have offered evidence that the lipide enzymatically produced from P-choline is lecithin. Their identification was based on the isolation of a highly purified radioactive lecithin by chromatography on alumina. These results were confirmed in the present investigation. Since P-choline is incorporated as a unit, the possibility that the actual product might be the phosphorus-free lipide observed by Kennedy2 to chromatograph with lecithin was excluded. Nevertheless, it was felt desirable to obtain additional proof that the product derived from P-choline was in fact lecithin. This was provided by the isolation of a water-soluble derivative, L-a-glycerophosphorylcholine, from the labeled lipide after mild hydrolysis by the method of Dawson (14). In this procedure, after selective hydrolysis of fatty acids, the water-soluble derivatives of the glycerophosphatides are separated by chromatography on paper. The labeled lipide yielded a single radioactive spot, RF = 0.84 in the phenol-ammonia solvent of Dawson (14), identical with the RF of an authentic sample of L-a-glycerophos-phorylcholine and closely similar to the value of 0.88 reported by Dawson.
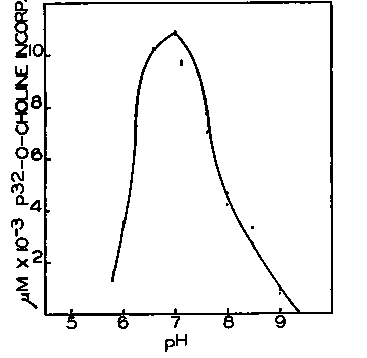
Fig. 1.
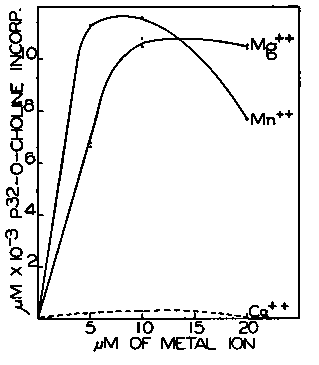
Fig. 2.
Fig. 1. Incorporation of P-choline-P" at various pH values. Each tube contained 20 pmoles of MgClj, 5 umoles of P-choline-P32 (109,000 c.p.m. per micromole), 10 pinoles of ATP (lot No. 116), 1.0 ml. of rat liver particles, and 100 pmoles of an equimolar mixture of phosphate and Tris. The final volume of the system was 2.0 ml. The pH of the Tris-phosphate buffer was varied to give the pH values shown, which were measured with a glass electrode after the addition of all reaction components. The tubes were incubated for 1 hour at 37°.
Fig. 2. The conditions of the enzyme assay were identical with those shown in Fig. 1, except that phosphate buffer of pH 7.4 was used and the added divalent cation concentration varied as indicated.
Optimal pH for Incorporation of P-choline into Lecithin-When the pH of the enzymatic reaction mixture was varied over a considerable range (Fig. 1), the highest incorporation of P-choline took place in the range 6.5 to 7.5. The activity of the enzyme system fell off rapidly at pH values below 6 and above 8.
Requirement for Divalent Cation-The enzymatic conversion of P-choline to lecithin requires the presence of added divalent cations. Manganese is somewhat more effective than magnesium at low concentration in this system, but at higher concentrations it becomes inhibitory (Fig. 2). Calcium is unable to activate the enzyme system; in fact, low concentrations (0.001 m) of calcium ion are inhibitory, even in the presence of optimal amounts of magnesium ion. A similar effect of calcium and barium ions has been reported by Rodbell and Hanahan (4).
Table II. Requirement Of Amorphous Atp For Incorporation Of P-Choline-P32 Into Lecithin
Vessel No. | Energy source | P-choline incorporated into lecithin |
mµmoles | ||
1 | Amorphous ATP | 20.6 |
2 | Succinate + AMP | 0.3 |
Each vessel contained 20 µmoles of MgCl2, 100 pmoles of phosphate buffer of pH 7.4, 10 pinoles of P-choline-P32 (27,500 c.p.m. per micromole), and 1.0 ml. of a suspension of freshly prepared rat liver particles. In the vessels containing amorphous ATP, 10 µmoles of ATP (lot No. 116) were added at the start of the experiment, and 1.0 µmoles every 10 minutes during the course of the reaction. To the other vessels, 100 pmoles of sodium succinate + 2 pmoles of AMP were added. The final volume of the system was 2.0 ml. The reaction was carried out in 20 ml. beakers, which were shaken for 1 hour at 37° in a Dubnoff apparatus.
Cofactor Requirement for Conversion of P-choline to Lecithin-No enzymatic conversion of P-choline to lecithin could be observed in experiments with rat liver particles unless ATP (amorphous preparation, Pabst lot No. 116) was added as cofactor. When it was attempted to replace the added ATP with crystalline AMP and an oxidizable substrate (succinate), which could then generate ATP by coupled oxidative phosphorylation in situ, no incorporation of P-choline to lecithin took place (Table II). This result was surprising, since earlier work (10) had indicated that coupled oxidative phosphorylation is effective in maintaining a source of metabolic energy for the incorporation of P32 into phosphatidic acids. It could be demonstrated in control experiments that the oxidative phosphorylation of AMP to ATP was indeed taking place in these experiments.
These results indicated that the actual reaction mechanism by which P-choline is converted to lecithin must be more complex than indicated in Reaction 1.
Role of CTP in Conversion of P-choline to Lecithin-When a crystallized preparation of ATP (lot No. 122) was substituted for the amorphous ATP used in the earlier experiments, no conversion of P-choline to lecithin could be detected. The identity and purity of the ATP lot No. 122 were established by a number of chromatographic, analytical, and enzymatic tests. It was also shown that the ATP lot No. 122 did not contain inhibitors of the enzyme system. It was therefore concluded that some cofactor other than ATP, present in trace amounts in the amorphous ATP lot No. 116, was necessary for the conversion of P-choline to lecithin.
Table III. Cofactor Requirement For Incorporation Of P-Choline-P32 Into Lecithin
Tube No. | Cofactor added | P-choline incorporated into lecithin |
mµmoles | ||
1 | 5 µmoles ATP (Lot 116) | 5.1 |
2 | 6 " 122) | 0.2 |
3 | 6 " 122) + 0.5 µmoles ITP | 0.0 |
4 | 5 '* 122) + 0.5 " UTP | 0.4 |
5 | 5 " " " 122) + 0.5 " GDP | 0.4 |
6 | 6 " 122) + 0.5 " CTP | 15.0 |
7 | 5 " 122) + 0.5 " " + 2.5 pmoles | 6.8 |
P-O-P | ||
8 | 5 pmoles ATP (Lot 122) + 0.1 pmole CTP | 10.0 |
9 | 6 " 122) + 0.05 " | 6.9 |
10 | 6 " 122) + 0.1 " CMP | 1.1 |
11 | 0.5 µmoles CTP (no ATP) | 8.0 |
Each tube contained 10 pmoles of MgCl2, 50 pmoles of phosphate buffer of pH 7.4,3 µmoles of P-choline-P" (113,500 c.p.m. per micromole), and 0.25 ml. of a 10 per cent suspension of lyophilized rat liver particles in a final volume of 1.0 ml. Other cofactors were added as indicated. The tubes were incubated for 1 hour at 37°.
Continue to:
My Books

