Formation Of Carbohydrate From Protein
Description
This section is from the book "Chemistry Of Food And Nutrition", by Henry C. Sherman. Also available from Amazon: Chemistry of food and nutrition.
Formation Of Carbohydrate From Protein
As early as 1876 Wolffberg tested the formation of carbohydrate from protein by fasting fowls for two days in order to free them from glycogen and then feeding for two days with meat powder which had been washed free from carbohydrate. Two of the fowls were killed soon after this protein feeding and showed more glycogen in their livers and muscles than could be accounted for except as derived from the protein fed. Two similar fowls killed 17 and 24 hours after feeding showed much less glycogen. This formation of glycogen from protein was fully confirmed by Kulz in a long series of experiments in which the food consisted of chopped meat thoroughly extracted with warm water (Lusk).
Independent evidence of the production of carbohydrates from protein is found in the work of Seegen, who chopped and mixed the liver of a freshly killed animal and determined the amount of carbohydrate in it by analysis of a portion, while the remainder was kept at body temperature and sampled for analysis from time to time. The percentage of carbohydrate was found to increase, showing that the liver cells can form car-bohydrate from their own protein substance.
The most striking evidence of the origin of carbohydrate from protein in the animal body is found in the many observations and experiments which have been made in cases of diabetes, and in experimental glycosuria produced either by administration of phlorizin or by removal of the pancreas. In such cases large amounts of carbohydrate may be given off in the form of glucose even when there is little body fat and no carbohydrate or fat is fed. The glucose must therefore result from the metabolism of protein. In Lusk's exhaustive experiments upon dogs rendered diabetic by phlorizin, 58 per cent of the total weight of protein broken down in the body (whether in fasting or on a meat diet) was eliminated in the form of glucose. According to Lusk: "After ingestion of protein in the normal organism this sugar becomes early available and may be burned before the nitrogen belonging to it is eliminated, or, if the sugar be formed in excess, it may be stored as glycogen in the liver and muscles for subsequent use. In this way it is obvious that at least half the energy in protein may be independent of the curve of nitrogen elimination, but may rather act as though it had been ingested in the form of carbohydrates."
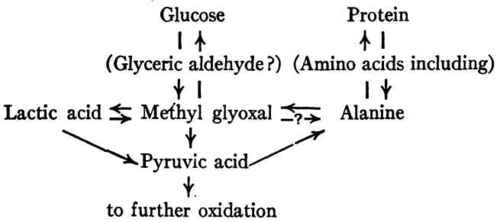
The way in which the production of carbohydrate from protein may take place has received much attention. Lusk demonstrated experimentally that alanine, one of the cleavage products of all known proteins, may yield glucose abundantly in the body; and he suggested that the change might occur through the formation of lactic acid as an intermediary product, since he had already shown that lactic acid is convertible into glucose. The work of Dakin has thrown further light on the intermediate steps of this transformation. He has shown that glyoxals have been formed from α-amino and α-hydroxy acids, in vitro - e.g. pyruvic aldehyde (methyl glyoxal) from alanine and lactic acid; and on the other hand α-hydroxy acids have been formed from glyoxals, both in vivo and in vitro.
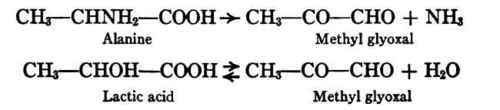
Attempts, however, to synthesize amino acids directly from glyoxals in vitro were not successful. There is some evidence of that synthesis in vivo, but it cannot be considered as fully established whether it takes place directly by the addition of ammonia to free glyoxals, or whether the α-amino acid is formed secondarily from the α-ketonic acid, resulting from the oxidation of glyoxals. The work of Knoop and of Embden and Schmitz leaves no doubt of the ability of the liver cells to form amino acids from the ammonium salts of the corresponding α-ketonic acids. Alanine, phenylalanine, and tyrosine were produced in this way.* It is of course possible that there may have occurred, in these liver perfusion experiments, intermediate steps not recognized by the investigators, but this does not detract from the significance of the fact that the synthesis of amino acids from ammonium salts has now been repeatedly demonstrated by experiment.
The relations emphasized by Dakin may be represented as follows:
* Embden also obtained alanine after perfusion of ammonium lactate, but the lactate may have been first changed to pyruvate and the alanine formed from the latter.
Attention may be called in passing to the possible importance of the interrelations of alanine, methyl glyoxal, and lactic acid to the regulation of neutrality, not only in the body as a whole (Chapter IX (Inorganic Foodstuffs And The Mineral Metabolism. The Elementary Composition Of The Body)) but also in the particular cells in which deamination may be more active than oxidation. It will be noted that alanine (a nearly neutral substance) yields on deamination another neutral substance (methyl glyoxal) and a base (ammonia)
CH3- CHNH2-COOH→ CH3-CO-CHO + NH3
And furthermore that the neutral substance methyl glyoxal may react with water to form lactic acid.
CH3-CO-CHO + H20 CH3-CHOH-COOH
Experiments in vitro have shown that the production of lactic acid from methyl glyoxal is promptly checked unless the free acid is quickly neutralized; also that the conversion of alanine into methyl glyoxal and ammonia is accelerated by acids (Dakin).
Thus far the possible mechanism of formation of carbohydrate from protein cleavage products has been considered here chiefly in terms of alanine. To what extent is its behavior representative of that of the other amino acids? Experiments in vitro show that the transformation of an α-amino acid into the corresponding α-ketonic aldehyde is a very general reaction. Dakin and Dudley demonstrated it for all the amino acids with which they worked - glycine, alanine, phenylalanine, valine, leucine, and aspartic acid. Experiments in vivo (chiefly on dogs rendered diabetic by phlorizin poisoning) have shown that glycine, alanine, serine, cystine, aspartic acid, glutamic acid, arginine, and proline are all capable of yielding large amounts of glucose. Leucine, tyrosine, and phenylalanine when similarly administered to phlorizinized dogs increase the elimination of acetoacetic acid rather than glucose. Valine, lysine, and tryptophane yield neither glucose nor acetoacetic acid to any important extent (Dakin).
The amino acids which yield glucose are called glucogenetic, and the amount of glucose which a given protein can yield in the body will naturally depend upon the glucogenetic amino acid radicles which it contains. Since the amino acids resulting from protein hydrolysis cannot be quantitatively recovered by any laboratory method thus far developed, it is not yet possible to calculate just how much carbohydrate a given protein should theoretically yield. For meat protein and some others the yield has been determined experimentally as in Lusk's investigations cited above. For further discussion of this point see Lusk's Science of Nutrition.
We have therefore abundant evidence from the work of independent investigators, using different methods, that the animal body may form carbohydrates readily and in large proportion from the protein of the food; and the mechanism of the process is beginning to be fairly well understood.
Continue to:
My Books

