Oxidation Of Carbohydrate. Continued
Description
This section is from the book "Chemistry Of Food And Nutrition", by Henry C. Sherman. Also available from Amazon: Chemistry of food and nutrition.
Oxidation Of Carbohydrate. Continued
Thus far in our study of the catabolism of glucose we have considered no oxidative changes but only the cleavages and transformations which, from the standpoint of the use of glucose as fuel, may be regarded as preliminary to oxidation. Probably the first oxidation product to be formed in glucose catabolism is pyruvic acid, CH3 • CO • COOH. This may be formed by the oxidation either of pyruvic aldehyde or of lactic acid. The relation of the three substances may be represented thus:
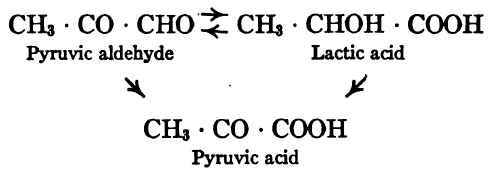
Pyruvic aldehyde and lactic acid are, so to speak, upon the same energy plane. Molecule for molecule they are of equal fuel value and either is readily convertible into the other. The conversion of pyruvic acid into lactic acid or pyruvic aldehyde probably takes place under certain conditions, but this involves reduction and so is not to be expected in the normal course of glucose oxidation. The fate of pyruvic acid under normal conditions is probably to undergo further oxidation through acetic acid to carbonic acid and water. It is possible that acetalde-hyde or alcohol or both may intervene between pyruvic acid and acetic acid, and that formic acid may be produced as an intermediate step between acetic and carbonic acids.
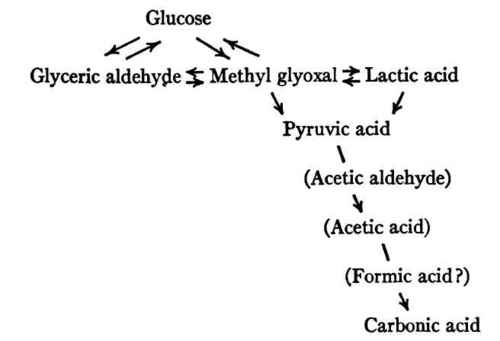
To summarize what now appears to be the most promising theory of the intermediary metabolism of carbohydrate, we may say that the glucose is first transformed, either directly or through glyceric aldehyde, into pyruvic aldehyde (methyl glyoxal), which may either be changed to lactic acid or oxidized directly to pyruvic acid that readily undergoes oxidation to carbon dioxide and water through steps not yet fully worked out. Lactic acid may also be converted into pyruvic acid and thus ultimately be completely oxidized. In case of excessive formation or inadequate oxidation, as in extreme muscular fatigue or asphyxial conditions, lactic acid may accumulate in the body or may be excreted unchanged.
Whatever the exact mechanism of the process, a large part of the glucose brought by the blood is oxidized in the muscles to furnish energy, which appears as external or internal work.
In general, the rate at which combustion takes place in the tissues depends upon the activity of the tissue cells, rather than upon the supply either of combustible matter or of oxygen. When a sufficient supply of oxygen is provided, any further increase has little effect upon the rate of combustion, and, as we have seen, any excess of carbohydrate instead of being burned is stored as glycogen. But while the absorption of an abundance of carbohydrate does not greatly change the amount of combustion taking place in the body, it may result in the use of carbohydrate as fuel almost to the exclusion of fat for the time being, as is shown by observations upon the respiratory quotient.
The respiratory quotient is the quotient obtained by dividing the volume of carbon dioxide given off in respiration by the volume of oxygen consumed. That is:
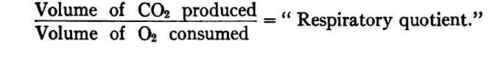
The numerical value of this quotient will evidently depend upon the elementary composition of the materials burned. Carbohydrates will yield a quotient of 1.0 since they contain hydrogen and oxygen in proportions to form water, so that all oxygen used to burn carbohydrate goes to the making of carbon dioxide, and each molecule of O2 so consumed will yield one molecule of CO2, occupying (under the same conditions of temperature and pressure) the same amount of space as the oxygen consumed to produce it. Thus in burning a molecule of glucose, six molecules of oxygen are consumed and six molecules of carbon dioxide produced:
C6H12O6 + 6O2->-6CO2 + 6H2O.
Here the volumes of oxygen and of carbon dioxide are equal and the respiratory quotient is 1.0.
Fats contain much more hydrogen than can be oxidized by the oxygen present in the molecule, and therefore a part of the oxygen used to burn fat goes to form water, so that the volume of oxygen consumed is greater than the volume of carbon dioxide produced, which gives a respiratory quotient lower than 1.0. The common fats of the body and of the food give quotients approximating 0.7. Thus the oxidation of stearin is represented by the equation:
2C57H110O6 + 163O2->114CO2 + 110H2O.
Since 163 volumes of oxygen are consumed and 114 volumes of carbon dioxide produced, the respiratory quotient is:
114 = 0.699. 163
Proteins give quotients intermediate between those of carbohydrates and fats, but if the amount of protein used in the body be determined by other methods (see Chapter VIII (Factors Determining The Protein Requirement)) and allowed for, one may then deduce from the respiratory quotient the proportions of carbohydrates and fats which are being burned in the body at any given time. The body will show a respiratory quotient of 1.0 when burning carbohydrate alone, of 0.7 when burning fat alone, and of an intermediate value when both fat and carbohydrate are being burned. If, now, the respiratory quotient rises soon after the eating of carbohydrate food, it is evident that the carbohydrate is being used more freely and fat less freely than before. .
In an experiment by Magnus-Levy the subject before taking food showed a quotient of 0.77. He then ate 155 grams of cane sugar, after which the quotient was determined at intervals of an hour for 7 hours with the following results: 1.01, 0.89, 0.89, 0.92, 0.82, 0.82, 0.79. The quotient here shows that within an hour after the sugar was eaten the body was making use of the carbohydrate to such an extent that fat either was not being used at all or was being formed from carbohydrate as fast as it was burned; and that for seven hours after the meal the body continued to use carbohydrate to a greater, and fat to a less, extent than was the case at the beginning of the Experiment.
It has been pointed out that, when carbohydrate is absorbed in larger quantity than is required to meet the body's immediate needs for fuel, the surplus normally accumulates as glycogen, which is stored conspicuously in the liver, but also to a considerable extent in the muscles and other organs. The amount' of carbohydrate which will be stored in the entire body after rest and liberal feeding is estimated at 300 to 400 grams. Thus the total amount of carbohydrate which can be stored as such in the body is no more than is frequently taken in one day's food.
When the supply of carbohydrate is so abundant that it continues in excess of the needs of the body and accumulates until the liver and muscles have no tendency to increase their store of glycogen, the further surplus of carbohydrate tends to be converted into fat.
Continue to:
My Books

