Protein-Sparing Action Of Carbohydrates And Fats. Continued
Description
This section is from the book "Chemistry Of Food And Nutrition", by Henry C. Sherman. Also available from Amazon: Chemistry of food and nutrition.
Protein-Sparing Action Of Carbohydrates And Fats. Continued
The food of the first period consisted of meat, milk, butter, bread, sugar, coffee, beer. That of the second period contained the same amounts of meat, milk, bread, coffee, and beer, but less sugar, more butter, and some bacon. The same amount of salt was taken in each case. The principal data of the experiment may be summarized as follows:
Nitrogen Balance When Feeding Isodynamic Quantities Of Carbohydrate And Fat (Tallquist)
Day | Intake | Output | |||||
Total nitrogen | Alcohol | Fuel value | Nitrogen | ||||
grams | grams | grams | grams | Calories | grams | ||
I | 16.27 | 44.0 | 466 | 18.5 | 2867 | 17.11 | - 0.84 |
2 | 16.27 | 44.0 | 466 | 18.5 | 2867 | 14.40 | + 1.86 |
3 | 16.27 | 44.0 | 466 | 18.5 | 2867 | 14.65 | + 1.62 |
4 | 16.27 | 44.0 | 466 | 18.5 | 2867 | 15.58 | + 0.69 |
5 | 16.08 | 140.0 | 250 | 19.0 | 2873 | 17.66 | - 1.58 |
6 | 16.08 | 140.0 | 250 | 19.0 | 2873 | 17.32 | - 1.24 |
7 | 16.08 | 140.0 | 250 | 19.0 | 2873 | 15.94 | + 0.14 |
8 | 16.08 | 140.0 | 250 | 19.0 | 2873 | 16.22 | - 0.14 |
Here only a part of the carbohydrate, about half of that present, and an amount representing about one third of the total fuel value of the diet, was replaced by fat. The change evidently had an unfavorable influence upon the nitrogen balance but the loss of body protein was relatively small and continued only 2 days.
Atwaterl compared the protein-sparing action of carbohydrate and fat in experiments in which the subject, an athletic young man of 76 kilos, performed a considerable amount of work. The experiments were carried out in the respiration calorimeter and covered in all 15 experimental days upon a diet rich in carbohydrates, arranged in four periods which were alternated with four equal periods in which the diet was rich in fats. The change from carbohydrate to fat and vice versa involved about 2000 Calories or nearly half the fuel value of the diet. The average results per day for the entire series of experiments were as follows:
On Diet Rich in Carbohydrates | On Diet Rich in Fat | |
Available Calories in food . . | 4532 | 4524 |
Heat equivalent of work performed, Calories..... | 558 | 554 |
Nitrogen in food, grams . . . | 17.5 | 17.1 |
Nitrogen in feces, grams . . . | 2.5 | 1.7 |
Nitrogen in urine, grams . . . | 16.6 | 18.1 |
Nitrogen balance, grams . . . | - 1.6 | - 2.7 |
Here again there is a difference in favor of the carbohydrate, but one which is so small as to be of almost no practical significance.
It appears that the carbohydrate of the food cannot be entirely replaced by an equal number of calories in the form of fat without an unfavorable effect upon the nitrogen balance; but that when the replacement is such as to affect not over one half of the total calories, the difference in protein-sparing action is but slight. Ordinarily on a normal mixed diet the same number of calories has about the same protein-sparing effect.
1 Ergebnisse der Physiologic, Vol. 3, Part I, page 497.
Landergren1 also found that it is only when the carbohydrate of the diet is entirely replaced by fat that the comparison is so strikingly against the fat as it seemed to be in Kayser's experiment. In Landergren's experiments the condition studied was not one of approximate equilibrium, but rather of nitrogen hunger. He fed men diets of adequate fuel value but containing only about one gram of nitrogen daily, and found that by four days of such feeding the urinary nitrogen may be reduced to about 4 grams per day. In one experiment in which the daily food contained 750 grams of carbohydrates the urine of the fourth day showed 3.76 grams of nitrogen. The carbohydrate was then entirely replaced by fat, with the result that the following days' urines contained respectively 4.28, 8.86, and 9.64 grams of nitrogen. Evidently in the case of a man accustomed to feeding largely upon carbohydrates the complete replacement of carbohydrate by fat leads to a loss (or an increased loss) of body protein. But by subsequent experiments of the same series it was found that a diet containing nearly half its calories in carbohydrate, and nearly half in fat, had 'apparently the same protein-sparing power as one made up almost exclusively of carbohydrates.
The explanation offered by Landergren is that when the diet supplies no carbohydrate, the glycogen of the body soon becomes exhausted, and the carbohydrate needed to keep up the constant glucose content of the blood is obtained largely by the breaking down of proteins.
This might suffice to explain the difference in effect of carbohydrate and fat, but not the fact that addition of a non-nitrogenous nutrient to a diet already sufficient may cause storage of nitrogen in the body.*
1 Skandinavisches Archiv fur Physiologie, Vol. 14, page 112 (1903); Abstract Experiment Station Record, Vol. 14, page 1099.
A satisfactory explanation of both sets of facts appears to be afforded by the recent advances in our knowledge of the fate of foodstuffs in metabolism which were outlined in Chapter V (The Fate Of The Foodstuffs In Metabolism Carbohydrates. Oxidation Of Carbohydrate). The outstanding relationships of the three groups of foodstuffs in the intermediary metabolism may be indicated schematically as follows:
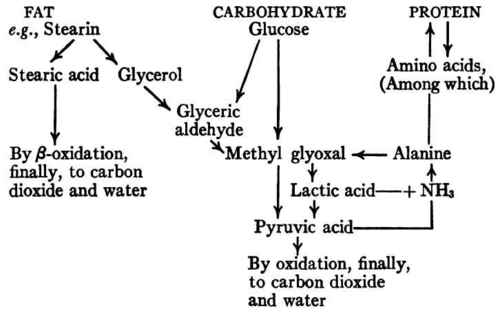
Since ammonia is always being formed in protein catabolism (by deaminization of amino acids), and since the ammonium salts of α-ketonic acids, such as pyruvic acid, are convertible into amino acids which are building materials for body protein, we have here a mechanism by which an intermediary product of carbohydrate metabolism (pyruvic acid) takes up a "waste product" of protein metabolism (ammonia) and turns it back into amino acid again. Thus carbohydrate, in undergoing metabolism, "spares" protein, not only by serving as fuel so that protein need not be drawn upon for this purpose, but also by furnishing material which in combination with ammonia (otherwise a waste product) can actually be converted in the body into some of the amino acids of which body proteins are composed and with which they are in equilibrium. This explains how an increased intake of carbohydrate, with resulting increase of pyruvic acid, naturally leads to increased synthesis of amino acids and thus to a tendency toward protein storage, or, to express the same thing in somewhat different terms, tends to push the reaction, Amino acidsDProtein, toward the right.
* Furthermore Lusk points out that Landergren's explanation is hardly adequate to cover the results obtained in gelatin-feeding experiments.
According to present theory, most, if not all, of the energy of the carbohydrate becomes available through oxidation processes which involve the intermediate production of pyruvic acid, an α-ketonic acid whose ammonium salt is capable of conversion into amino acid. Of the fat only the glyceryl radicle (about one twentieth of the fuel value) is oxidized through pyruvic acid, while the fatty acid radicles, representing about nineteen twentieths of the energy of the fat, are metabolized through ß-oxidation processes which yield, so far as we know, no product whose ammonium salt is convertible into amino acid in the body. Hence complete withdrawal of carbohydrate, even though substituted by sufficient fat to yield an equal number of calories, must be expected to result in increased excretion of nitrogen; but when no more than half of the carbohydrate is replaced by fat there seems to be enough pyruvic acid produced to meet the practical requirements of economical metabolism of protein.
Continue to:
My Books

