Mechanisms Of Hormone Action. Part 7
Description
This section is from the book "The Scientific Contributions Of The Ben May Laboratory For Cancer Research", by The University of Chicago. Also available from Amazon: The Scientific Contributions Of The Ben May Laboratory For Cancer Research.
Mechanisms Of Hormone Action. Part 7
ii. Hydrogen transport between nucleotides by 3a-hydroxysteroid dehydrogenase. With knowledge of the pyridine nucleotide and analog specificity of this dehydrogenase, it was possible (111) to demonstrate the following reactions in the presence of catalytic amounts of 3a-hydroxy- or 3-ketosteroids (Table VI):
(i) DPNH + acetylpyridine-DPN + -→ DPN+ + acetylpyridine-DPNH
(ii) DPNH+ pyridine aldehyde-DPN → DPN + + pyridine aldehyde-DPNH
(iii) DPNH + thimcotinamide-DPN+ → DPN++ thionicotinamide-DPNH
Because of differences in absorption spectra between DPNH and the reduced analogs of DPN, these reactions could be measured directly by differential spectrophotometry. The enzymatic hydrogen transfer to the DPN-analogs was dependent upon the presence of small amounts of appropriate steroids and was proportional to enzyme concentration. The dependence of reaction (iii) on steroid and enzyme is shown in Fig. 4. The reaction with 8.6 ug. of purified enzyme was completely steroid dependent, whereas with 43 ug. of enzyme there was a slow transfer of hydrogen in the absence of added steroid which may be attributed to the presence of minute amounts of steroid in the enzyme preparation, or more probably, to contamination of the enzyme by extraneous flavoproteins which transfer hydrogen to analogs of higher potential. Androsterone and androstane-3, 17-dione are equally effective in promoting hydrogen transport between DPNH and DPN analogs. A significant rate of hydrogen transfer in reactions (ii) and (iii) is catalyzed by (3.5 to 7.0) X 10-6 M steroid. The hydrogen transfer between DPNH and the thionicotinamide analog of DPN has an optimum at pH 8.5 to 8.8 It may be calculated from the data presented in Fig. 4 that 35 mumoles of androstane-3,17-dione transported about 100 mumoles of hydrogen per minute at 25° C. in the presence of 43 ug. of enzyme. The same quantity of enzyme under similar conditions would have reduced about 1200 mumoles per minute of DPN and 800 mumoles per minute of the thionicotinamide analog of DPN. Hence the rate of transhydrogenation is about 10% of the rate of dehydrogenation in this system.
Table VI. Hydrogen Transfer Between Dpnh And Analogs Of Dpn Catalyzed By Bacterial 3a.-Hydroxysteroid Dehydrogenase*
System | Amount of enzyme (ug.) | DPNH (µmoles) | Acceptor nucleotide (µmoles) | Rate of reduction of acceptor nucleotide (µmoles/min./ mg. protein) |
DPNH → thio- | 8.6 | 0.5 | 0,30 | 1.51 |
nicotinamide-DPN | 8.6 | 0.5 | 0.61 | 2.27 |
8.6 | 0.5 | 1.22 | 3.07 | |
8.6 | 0.5 | 2.44 | 3.05 | |
Measured at 400 run (E | = 7150) | |||
DPNH → acetyl- | 21.5 | 0.5 | 2.38 | 0.0695 |
pyridine-DPN | ||||
Measured at 400 mµ (E | = 2500) | |||
DPNH → pyridine | 43 | 0.5 | 0.625 | 0.134 |
aldehyde-DPN | 43 | 0.5 | 1.25 | 0.291 |
43 | 0.5 | 3.75 | 0.438 | |
Measured at 385 mµ(E | = 4130) | |||
* Velocities were measured at 25° C. in systems of 3.0 ml. final volume containing: 200 (imoles Tris of pH 8.5; 10 µg. androstane-3,17-dione in 0.01 ml. methanol; enzyme, DPNH, and acceptor nucleotides in amounts indicated. The enzyme had a specific activity of 83,700 units per milligram protein. Each velocity determination involved the use of three cuvettes. The control contained no nucleotide and no steroid, a second cuvette contained all ingredients except the steroid, whereas the third cuvette contained the complete system. The rates given are corrected for the small velocity of reduction of acceptor nucleotide in the absence of steroid.
The rate of hydrogen transport between donor and acceptor nucleotides depends quite critically upon the relative concentrations of these reactants. This relation has been examined for the hydrogen transfer from DPNH to the thionicotinamide analog of DPN by purified bacterial 3a-hydroxysteroid dehydrogenase in the presence of catalytic quantities of androstane-3, 17-dione. Figure 5 shows the velocity of reduction of the thionicotinamide analog of DPN for two concentrations of the analog (1.2 X 10-4 M and 4.1 X 10-4 M) and a range of DPNH concentrations (3.3 X 10-5M to 3.3 X 10-4). The optimum ratio of acceptor to donor nucleotides is about 2 to 3, and the reaction velocity declines markedly for higher or lower ratios. This finding indicates that the site(s) on the enzyme to which the nucleotides become attached are the same or interact with one another. If the transfer of hydrogen depends upon these sites alternately binding the reduced donor and oxidized acceptor nucleotides (and thereby effecting oxidoreduction of steroid), then a competition between donor and acceptor nucleotides of this type is to be expected.
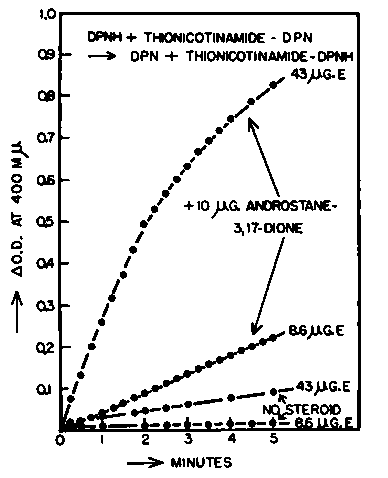
Fig. 4. Hydrogen transfer from DPNH to the thionicotinamide analog of DPN catalyzed by purified bacterial 3a-hydroxysteroid dehydrogenase in the presence of androstane-3,17-dione.
The reactions were carried out at 25° C. in cuvettes of 1.0 cm. light path. The systems contained in a final volume of 3.0 ml.: 200 umoles Tris buffer of pH 8.5, 0.5 umole DPNH, 0.73 (imole of the thionicotinamide analog of DPN, 10 ug. of androstane-3,17-dione in 0.01 ml. methanol or 0.01 ml. methanol alone, and 8.6 ug. (720 units) or 43 µg. (3600 units) of purified bacteria] 3a-hydroxysteroid dehydrogenase. The optical density was measured at 400 mµ, against a control containing no DPNH and no steroid.
b. Bacterial β-Hydroxysteroid Dehydrogenase, i. Reactivity of β-hydroxysteroid dehydrogenase with DPN analogs. Purified preparations of bacterial β-hydroxysteroid dehydrogenase (83) reduced DPN, the acetyl-pyridine and pyridine aldehyde analogs of DPN, but neither TPN nor deamino-DPN, in the presence of substrate concentrations of testosterone or of estradiol-17β. The relative velocities of reaction of the two analogs of DPN at saturating concentrations and pH 9.5 were about one-fifth to one-third the reaction rate with DPN using estradiol-17β as substrate.
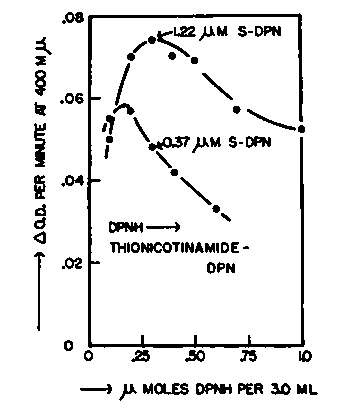
Fig. 5. The rate of hydrogen transfer from DPNH to the thionicotinamide analog of DPN catalyzed by purified bacterial 3a-hydroxysteroid dehydrogenase with androstane-3,17-dione as carrier. The graph illustrates the influence of the relative concentrations of donor and acceptor nucleotides.
The reactions were carried out at 25 C. in cuvettes of 1.0 cm. light path. The systems contained in a final volume of 3.0 ml.: 300 umoles of Tris buffer pH 8.5, varying quantities of DPNH, 0.37 or 1.22 umoles of the thionicotinamide analog of DPN (S-DPN), 8.6 ug. (720 units) of purified bacterial 3a-hydroxysteroid dehydrogenase, and 10 ug. androstane-3,17-dione in 0.03 ml. methanol. The optical density at 400 mµ was measured at intervals, and the initial linear rates were expressed as changes in optical density per minute.
ii. Hydrogen transport by β-hydroxysteroid dehydrogenase. A steroid-dependent transfer of hydrogen between DPNH and certain analogs of DPN was demonstrated with this enzyme (111) in the presence of catalytic levels of 17P-hydroxy- and 17-ketosteroids, as follows:
(iv) DPNH + acetylpyridine-DPN + → DPN++ acetylpyridine-DPNH
(v) DPNH + pyridine aldehyde-DPN + -→ DPN + + pyridine aldehyde-DPNH
With purified preparations of 8-hydroxysteroid dehydrogenase, only low rates of hydrogen transfer occurred in reaction (v) in the absence of added steroid. However, a variable steroid-independent blank was observed in reaction (iv) with different enzyme preparations. The steroid-independent component of these reactions was probably catalyzed by contaminating flavoproteins which transfer hydrogen between reduced pyridine nucleotides and analogs of higher oxidoreduction potential rather than by bound steroids contained in the enzyme preparations. The hydrogen transfer from DPNH to the pyridine aldehyde analog of DPN was studied by differential spectrophotometry at 385 mu. Table VII gives the rates of this reaction as function of the acceptor nucleotide concentration. The pH optimum is about 8.7 to 8.9. Most of the studies have been carried out with about (1-2) X 10-5M 4-androstene-3,17-dione as carrier. Half maximum velocity was obtained with a steroid concentration of somewhat less than 3 X 10-6M (about 1 ug. per milliliter) (Fig. 6).
Table VII. Effect Of Nucleotide Acceptor Concentration On The Hydrogen Transfer From Dpnh To The Pyridine Aldehyde Analog Of Dpn Catalyzed By Bacterial β-Hydroxy Steroid Dehydrogenase*
Continue to:
My Books

